
One of the ideas that have surely crossed the mind of every beekeeper is that of importing other more productive subspecies of bees. Or at least it is said that they are more productive, since we are all aware that the rumor mill in our sector is the order of the day. However, at least in the Iberian Peninsula, few technicians recommend these practices, since our bees are especially well adapted to our environment.
In relation to this topic there is a question that always comes to mind: how did the current subspecies of bees originate?
About the origin of our bee
The Apis genus is naturally present in many areas: Europe, Asia and Africa. Within this genus three groups are distinguished by their genetic characteristics: cavity nesting bees (A. mellifera, A. cerana and A. koschevnikovi), giant bees (A. dorsata and A. nigrocincta) and dwarf bees (A. florea and A. andreniformis) [1,2] . All are found in Asia except A. mellifera.
29 subspecies of A. mellifera have been described based on their morphology [3], which split from each other at least 6 million years ago. We can group them into five large groups [4,5]:
- group A, which includes most of the African subspecies
- group M, which includes the central and northern European subspecies (ours)
- group C, which includes the Eastern European subspecies
- group O, which includes those from the Middle East
- and group Y, which is limited to the area of the Horn of Africa
The most widely accepted hypothesis today for the origin of our bees was prop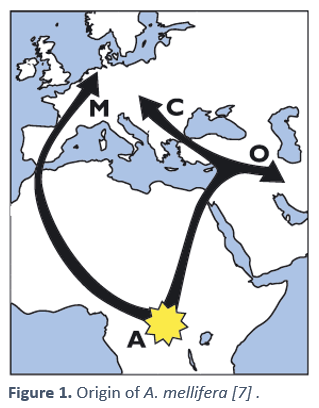 osed by EO Wilson [6], based on their different abilities to form a winter cluster. Wilson argued that this behavior would be an adaptation of bees to temperate climates, since it does not occur in Asian bee populations. European bees would most likely come directly from Africa through the Strait of Gibraltar. [7]. This hypothesis would be reinforced by an important genetic study carried out by Whitfield et al. in 2006 [8], in which it was seen that the bees of the M and C lineages were very different, despite both being European, with M being more similar to A, and C to O.
osed by EO Wilson [6], based on their different abilities to form a winter cluster. Wilson argued that this behavior would be an adaptation of bees to temperate climates, since it does not occur in Asian bee populations. European bees would most likely come directly from Africa through the Strait of Gibraltar. [7]. This hypothesis would be reinforced by an important genetic study carried out by Whitfield et al. in 2006 [8], in which it was seen that the bees of the M and C lineages were very different, despite both being European, with M being more similar to A, and C to O.
Other behaviors differ greatly between European and African bees, such as migratory behavior, reproductive rates and defensive behavior. So then, did bees acquire new behaviors as they colonized Europe, or did the populations from which they originated already have these adaptations innately, even if they did not put them into practice?
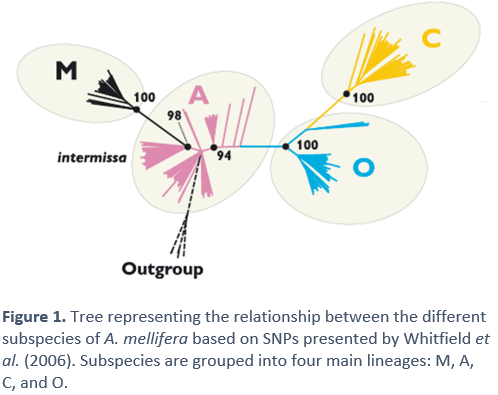 Genetic studies tell us that the genetic variation between some bee populations and others are relatively low, which indicates that the subspecies of A. mellifera have not experienced long periods of isolation. The studies by Whitfield et al. [8], were able to clearly delimit the subspecies and groups, within those that had already been previously defined by morphological studies (see figure 2). However, only one subspecies appeared to be a mixture of groups A and M: A.m. intermissa. This could indicate that this subspecies has a recent hybrid origin between A and M, which would support the thesis that A. mellifera reached Western Europe through the Strait of Gibraltar. In addition, the recreation of phylogenetic trees based on DNA variability also seems to show that there is a closer relationship between sub-Saharan and Western European bees than between the latter and Eastern European bees.
Genetic studies tell us that the genetic variation between some bee populations and others are relatively low, which indicates that the subspecies of A. mellifera have not experienced long periods of isolation. The studies by Whitfield et al. [8], were able to clearly delimit the subspecies and groups, within those that had already been previously defined by morphological studies (see figure 2). However, only one subspecies appeared to be a mixture of groups A and M: A.m. intermissa. This could indicate that this subspecies has a recent hybrid origin between A and M, which would support the thesis that A. mellifera reached Western Europe through the Strait of Gibraltar. In addition, the recreation of phylogenetic trees based on DNA variability also seems to show that there is a closer relationship between sub-Saharan and Western European bees than between the latter and Eastern European bees.
If we take into account the morphological and behavioral characteristics of the different subspecies of bees, these major differences between the bees of Eastern and Western Europe are not appreciated. This could be because the populations of bees, which were genetically quite different when they arrived in Europe from both sides of the Mediterranean, found a similar climate, so they ended up developing similar adaptations.
In African populations there is greater genetic variability, which could indicate that in that area bees have been developing and evolving for a longer time; however, there are authors who argue that this is so because during the quaternary ice age, bee populations in Europe and Asia were considerably reduced, thus also reducing their genetic variability, while in Africa they were able to continue to evolve more normally.
Consequences of the modern movement of the domestic bee
As we have already seen, one of the greatest successes of A. mellifera is undoubtedly its ability to adapt. These social insects have managed, with help from humans, to colonize a large part of our planet's landmasses. But how far do the consequences of this adaptive success go? What about native pollinators? And with the other subspecies of A. mellifera present in certain areas? The domestic bee managed to spread throughout Africa and Eurasia naturally, but could the movements produced by humans pose a significant risk to the survival of other races of bees and native pollinators? Could A. mellifera be considered an invasive species?
According to the definition of Lonsdale [9], an invasive species should have three characteristics:
- They proliferate and spread through the environment.
- They are destructive to biodiversity or to human interests.
- They have moved from one geographic area to another where they were not previously present.
Although points 1 and 3 are clearly applicable to our bees, in 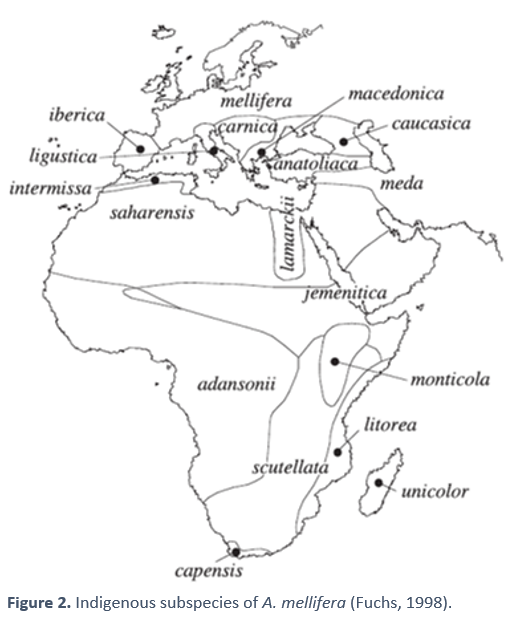 the case of point 2, it is not so clear. To analyze these possible consequences, we are going to divide the casuistry into three different situations:
the case of point 2, it is not so clear. To analyze these possible consequences, we are going to divide the casuistry into three different situations:
- regions where A. mellifera is endemic
- regions where A. mellifera is not naturally present, but other Apisspecies are present
- and regions in which there are no endemic species of the genus Apis.
Given the very high intensity and magnitude of the global bee trade, the few cases of A. mellifera invasions with a clear negative impact on the environment are surprising.
What is less surprising is that, owing to this bee trade, bee diseases and parasites have spread globally. [10]. This is precisely one of the main problems of the introduction of bee populations in areas where they were already present, as is the case of the expansion of varroa, Varroa jacobsoni initially, and now V. destructor, from Asian to European bees [11]. And probably that of Nosema ceranae, also from Asia to Europe, although it could have been introduced through the honey trade and its use as food for bees. A more complex case is that of China, where the populations of A. cerana have dropped considerably, since the queens of A. cerana and the drones of A. mellifera mate, giving rise to infertile individuals [12].
There is also a risk of loss of local subspecies. And the fact is that, due to the particular livestock system of beekeeping, which allows populations introduced by humans to share their genetic material with wild populations, these may be at risk of disappearing. Indeed, the massive introduction of populations of highly selected bees for beekeeping production may cause certain adaptive characteristics of local bees to be diluted, losing a genetic wealth that may be of interest with regard to the future. This is especially important in an environment as changeable and unpredictable as the current one.
In the case of the Americas, there were no bees of the Apis genus naturally, so colonies were imported from Europe around 300 years ago, with swarms escaping and establishing themselves in nature [13]. These bees did not adapt well to subtropical climates, so, in 1956, experts from Brazil introduced tropical African bees to improve honey production in their areas with that climate [14]. Specifically, they chose the scutellatasubspecies.
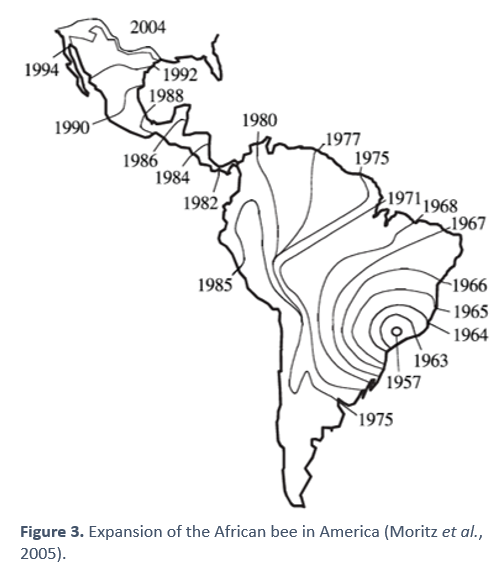 These bees quickly hybridized with the Brazilian ones (of European origin), and then with those of neighboring and nearby countries (also European). The "Africanized" ones progressively invaded a large part of the continent (see figure 4), giving rise to a very important problem in beekeeping management in the region [15].
These bees quickly hybridized with the Brazilian ones (of European origin), and then with those of neighboring and nearby countries (also European). The "Africanized" ones progressively invaded a large part of the continent (see figure 4), giving rise to a very important problem in beekeeping management in the region [15].
The adaptive success of hybrids of A. m. scutellata over the European ones is due to a series of factors: greater number of swarms per colony and their greater mobility, greater production of drones, smaller colonies, shorter queen development time, ability to usurp other colonies, greater aggressiveness, greater ability to diversify food resources, seasonal migration and less storage of honey for the winter [16]. All this makes "Africanized" bees superior to European bees in their ability to adapt to tropical climates.
The promotion of local subspecies as an option for the future
As we have seen, since its origin, A. mellifera has been able to adapt its reproductive and feeding strategies, which has allowed it to colonize many climates and areas, from that origin in Africa.
In addition, they have managed to adapt very well to their livestock use by humans, which has definitively led to their spreading around practically the entire globe.
However, we must not ignore the problems that can be caused by the movement of bee populations from one place to another. We have already suffered disasters at health level (with the globalization of certain diseases), population, production, etc., and we cannot rule out the possibility of causing environmental problems in the future.
For all these reasons, we must get used to working with local bees, selecting them to promote the characteristics that interest us and to reduce those that are not, while avoiding new imports of exotic bees that could put our beekeeping and our ecosystem at risk.
Bibliographic references:
- Arias, M.C.; Sheppard, W.S. Molecular phylogenetics of honey bee subspecies (Apis mellifera L) inferred from mitochondrial DNA sequence. Molecular Phylogenetics and Evolution 1996, 5, 557-566, doi:10.1006/mpev.1996.0050.
- Raffiudin, R.; Crozier, R.H. Phylogenetic analysis of honey bee behavioral evolution. Molecular Phylogenetics and Evolution 2007, 43, 543-552, doi:10.1016/j.ympev.2006.10.013.
- Sheppard, W.S.; Meixner, M.D. Apis mellifera pomonella, a new honey bee subspecies from Central Asia. Apidologie 2003, 34, 367-375, doi:10.1051/apido:2003037.
- Garnery, L.; Cornuet, J.M.; Solignac, M. Evolutionary history of the honey bee Apis mellifera inferred from mitochondrial DNA analysis. Molecular Ecology 1992, 1, 145-154, doi:10.1111/j.1365-294X.1992.tb00170.x.
- Franck, P.; Garnery, L.; Loiseau, A.; Oldroyd, B.P.; Hepburn, H.R.; Solignac, M.; Cornuet, J.M. Genetic diversity of the honeybee in Africa: microsatellite and mitochondrial data. Heredity 2001, 86, 420-430, doi:10.1046/j.1365-2540.2001.00842.x.
- Wilson, E.O. The insect societies; Cambridge, Massachusetts, USA, Harvard University Press [Distributed by …: 1971.
- Han, F.; Wallberg, A.; Webster, M.T. From where did the Western honeybee (Apis mellifera) originate? Ecology and Evolution 2012, 2, 1949-1957, doi:10.1002/ece3.312.
- Whitfield, C.W.; Behura, S.K.; Berlocher, S.H.; Clark, A.G.; Johnston, J.S.; Sheppard, W.S.; Smith, D.R.; Suarez, A.V.; Weaver, D.; Tsutsui, N.D. Thrice out of Africa: Ancient and recent expansions of the honey bee, Apis mellifera. Science 2006, 314, 642-645, doi:10.1126/science.1132772.
- Lonsdale, M. A global perspective on biological invasions–impacts on ecosystems and economics. Keynote address. In Proceedings of the Proceedings of the World Conservation Congress, Bangkok, 2004; pp. 17-25.
- Page, R.; Guzmán-Novoa, E.; Morse, R.; Flottum, K. The genetic basis of disease resistance. Honey Bee Pests, Predators, and Diseases. Medina OH: AI Root Co 1997, 469-492.
- Anderson, D.L.; Trueman, J.W.H. Varroa jacobsoni (Acari : Varroidae) is more than one species. Experimental and Applied Acarology 2000, 24, 165-189, doi:10.1023/a:1006456720416.
- Li, W. Introducing Apis mellifera threatens Apis cerana. Journal of Bee 1998, 23, 4-6.
- Spivak, M.; Fletcher, D.J.; Breed, M.D. The African honey bee; CRC Press: 2019.
- Kerr, W.E. The history of the introduction of African bees in Brazil. South African Bee J. 1967, 39, 33-35.
- Schneider, S.S.; Hoffman, G.D.; Smith, D.R. The African honey bee: Factors contributing to a successful biological invasion. Annual Review of Entomology 2004, 49, 351-376, doi:10.1146/annurev.ento.49.061802.123359.
- Moritz, R.F.A.; Hartel, S.; Neumann, P. Global invasions of the western honeybee (Apis mellifera) and the consequences for biodiversity. Ecoscience 2005, 12, 289-301, doi:10.2980/i1195-6860-12-3-289.1.



